
Montane systems
The montane environment differs from that at lower elevations , primarily because the climate is cooler.
Temperature is colder at higher elevations because rising air expands and expanding air cools. Precipitation in montane environments often comes in the form of snow. The growing season for plants is short, and decomposition (which is highest in moist, warm environments) is slow.
Snow can affect plants by burying plants for part of the year, by accumulating on branches and adding weight to those branches, and by damaging plant tissues through wind-blown snow and ice crystals.
Soils in the mountains are often thin and nutrient-poor. Thin soils occur because
There are areas of soil accumulation, such as in the bottoms of valleys and canyons in the mountains, where both soil and water accumulate and may cause mountain meadows to form.
Abrasion from blowing ice crystals can severely damage plant tissue and kill it.
Plants that are covered by a blanket of snow during winter are protected from this type of damage. In very high elevations, where vegetation is sparse and blowing ice crystals are a real hazard, most plants are short. Cushion plants and mat-like plants (below) have two advantages over taller plants:

At the upper limit of forests (tree line), wind-blown ice crystals damage and shape the trees. This vegetation is known as Krummholz vegetation. Abrasion by blowing snow and ice kills buds, either pruning the tree into a mat-like form or killing buds only on the windward side of the tree to produce a "flag" form.

Snow load can also damage plants. Trees in snowy areas tend to be flexible to shed snow without breaking. Some trees are winter deciduous, and present less surface area to catch snow. Others, however, like the dominant, evergreen gymnosperms, just have to be flexible. They typically have flexible branches and tough needles that can resist damage by ice crystals and droop to shed snow.

In California, heavy snow that extends to lower elevations than normal can severely tree species that are not adapted to it. For example, blue oaks and valley oaks of the foothill woodlands tend to break: their rigid branches do not shed snow easily.
Nutrient-poor conditions tend to select for plants who retain their leaves for a long time. This is because, although plants tend to withdraw a large fraction of the phosphorus, nitrogen, and other nutrients from their leaves before the leaves are dropped, they can never recover all of it. Therefore, every time a plant drops a leaf, it loses nutrients. In a nutrient-poor environment, it is difficult to replace those lost nutrients.
One way to conserve nutrients is to retain leaves for a long time, maximizing the amount of photosynthesis the plant gets out of its investment in individual leaves. It is thought that this is the main reason we see long-lived sclerophyllous leaves in a wide range of nutrient-poor environments (hard chaparral, bogs, montane forests, etc.).
Although there is a wide variety of plant types in montane systems, the nutrient poverty of soil may be the main reason we see forests dominated by coniferous forests with long-lived evergreen leaves. (Contrast eastern deciduous forests, which also experience snowy cold winters but grow on deeper, richer soil.) Some gymnosperms retain their needles only a few years, but some pines may retain their needles for decades.
There are many ways of dealing with a short growing season. Short perennials, or mat plants, may extend their growing season by sheltering in the boundary layer of the ground, which heats up earlier in the spring. (This was discussed above.) Annuals must complete their life cycle quickly, and many of the mountain annuals do not get large. In many species, the short growing season has selected for early flowering, ensuring that the plant produces some seed before the winter sets in. This is discussed on the next page.
Many species grow across a wide range of elevations. In most, the plants that grow at high elevation look different from those that grow at low elevation. In some cases, those differences may simply be due to the fact that they develop under different conditions and, therefore, develop differently. In other cases, the differences are genetically based.
If a species has populations grow in different environments, these populations (or sets of populations) are considered different ecotypes if
In terms of high- and low-elevation ecotypes, this means that populations at high elevation have genetically based traits that allow them to perform better at high elevation. Conversely, populations at low elevation have genetically based traits that allow them to perform better at low elevation. If you try taking seed from high elevation and planting it at low elevation, the progeny will not perform as well as progeny of the low-elevation population. The converse is also true.
In some cases, we recognize these differences taxonomically as subspecies or varieties of a species. In other cases, the genetic differences are functionally important, but may not be obvious enough to merit taxonomic distinction.
One species that has four recognized varieties is California buckwheat (Eriogonum fasciculatum). Most plants found on the CSUSB campus are Eriogonum fasciculatum var. foliosum. This variety can grow to be 60 - 150 cm tall. It does not grow above an elevation of 1300 m.
In contrast, the variety that reaches the highest elevation (2500 m), is a shorter variety: Eriogonum fasciculatum var. polifolium. This variety only grows to be 20-60 cm tall. 
Similarly, there are multiple varieties of Artemisia tridentata in California. The most common variety at lower elevations in transmontane regions of California is big sagebrush (or basin big sagebrush). It grows may grow to a height of 2 m, and it grows at elevations below 1900 m. Mountain sagebrush may grow to a height of 1 m, but it keeps its leaves low (see photo). It grows at elevations of 1800 - 3000 m and higher.

Check your understanding:
Remember what an ecotype is.
Check your understanding:
Investigators have been exploring the existence of ecotypes in plant species since the 1920s. The standard approach to identifying whether populations are genetically different has been to grow plants from different populations together in a common environment (e.g., from seed) and see if plants from different populations retain any differences that were seen in the field. Since the plants are being grown in a common environment, any differences seen are probably genetically based differences. The "common garden" experiment grows plants together in a common environment to identify genetically based differences.
To determine whether populations are adapted to local conditions further investigation is needed. Reciprocal transplants, growing plants from different populations each other's native environments, are used to test for local adaptation.
Check your understanding:
in the 1940s, Clausen, Keck and Hiesey, from the Carnegie Institution at Stanford University, set up extensive experiments that tested for ecotypic differences in a number of species across a range of elevations in California.

The figure below shows just one example of their results. The height of yarrow plants grown from seed collected from different populations and grown in a common environment differed. This figure does not show heights of plants growing in these different locations. It shows the height of plants grown in a common environment but collected from nine different locations. These locations spanned an elevational range of over 2000 m in the Sierra Nevada Mountains.
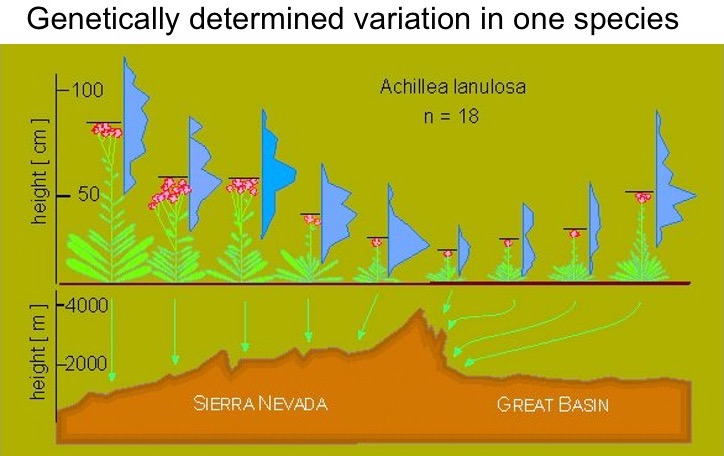
The pattern shown above, with populations from higher elevations being genetically short, was found repeatedly in multiple species.
Other traits that varied included traits that were not so obvious, such as the date that flowering began.
Fire plays an important role in shaping the structure of montane forests.
Fire tolerance varies among species. Some trees have thick bark and survive low-intensity ground fires. Other tree species have thinner bark and are killed, even by low-intensity ground fires.
Fire tolerance varies with age. Even tree species that develop thick bark as adults are vulnerable to fire during the first few decades of their life. Seedlings and saplings (young trees) have thinner trunks, thinner bark, and leaves that are closer to the ground where they are more likely to be consumed in a ground fire.
In many of California's montane forests, fire occurred more frequently than it has in recent decades, keeping the understory of the forest open and causing the fires that did occur to remain as low-intensity ground fires.

Giant sequoia (Sequioadendron giganteum) is a species that is particularly suited to a regime of low-intensity ground fires. Mature trees have very thick bark, so they survive ground fires. Additionally, the seeds require open mineral soil to germinate and establish seedlings. This means that seedlings only establish after leaf litter and duff (partially decomposed leaf litter) has burned off. Fire-suppression policies of the 1900s allowed encroachement of more fire sensitive species, such as firs (genus Abies) into the giant sequoia groves, providing "ladder fuels" that would carry flames up into the crowns of the giant sequoias, potentially killing them, and creating conditions that suppressed regeneration of the big trees. The photos below are from the National Park Service's archives.

It is important to remember that not all conifers are tolerant of fire. Piñon pines, which dominate forests on the dry, transmontane side of California's major mountain range, have thin bark and do not tolerate even low intensity ground fires.
|
From your previous lesson (Week 6) |
|
...pinyon pines do not tolerate fire well. Mature pinyon pine trees do not have bark that is thick enough to protect them, and even low-intensity ground fires will kill the trees. Researchers from UC Riverside have estimated that, historically, pinyon pine forests in southern California burned, on average, only once every 480 years. They also estimated that it took 100-150 years for a burned pinyon pine woodland to recover enough to look the way it did before a fire (Wangler and Minnich, 1996). As a result, using prescribed fire is probably not a good way to manage pinyon pine forests if the objective is to keep the pinyon pine forest healthy. |

Serotiny is a characteristic of many fire-adapted conifers. Serotinous cones stay on the tree for many years, and their cone scales do not separate to release their seeds until they are heated. This allows cones to release seeds right after fire. Seedlings then enjoy an environment that is open and fairly free from competition.
In most cases, trees with serotinous cones are not fire-tolerant. They die in fires and the persistence of the species on the site depends on post-fire regeneration from seedling establishment.

There are several pine species with serotinous cones in California. You can tell if a tree has serotinous cones by looking at the placement of the cones on the tree. If a tree drops its cones or disperses its seeds soon after they mature, you will only see mature seed cones near the ends of branches. If the cones remain on the tree for many years, delaying seed release (i.e, they are serotinous), you will see cones that are still attached to older sections of stems.

Three things to note about species with serotinous cones.
Check your understanding:

And consider: